CONCEPT · ENTRY 026 · R2 BIOLOGICAL
Bioelectric Governor
The tissue-scale dynamical control variable that governs morphogenetic targets independently of the genome — a gap-junction-coupled bioelectric attractor whose rewriting can redirect the tissue's construction plan while the parts list remains fixed.
- Register
- R2 biological — metabolic-closure.
- Burn rate
- ATP per ion-pump cycle; continuous, tissue-scale metabolic expenditure to maintain transmembrane voltage gradients
- Genealogy
- Levin 2012 · Levin & Martyniuk 2018 · Simondon 1958
- Appears in
- Chapter 8 — The Bioelectric Governor
- What it is not
-
- Not the genome or its molecular expression — the Bioelectric Governor is a dynamical control variable above the parts list.
- Not consciousness or sentience at the tissue level — operational normativity without interiority.
- Not a metaphor for governance — a literal, measurable, pharmacologically accessible control variable.
- Not sufficient for cognition — the bioelectric stratum is the first cognitive closure, not the whole of it.
DIAGRAM
The Bioelectric Governor
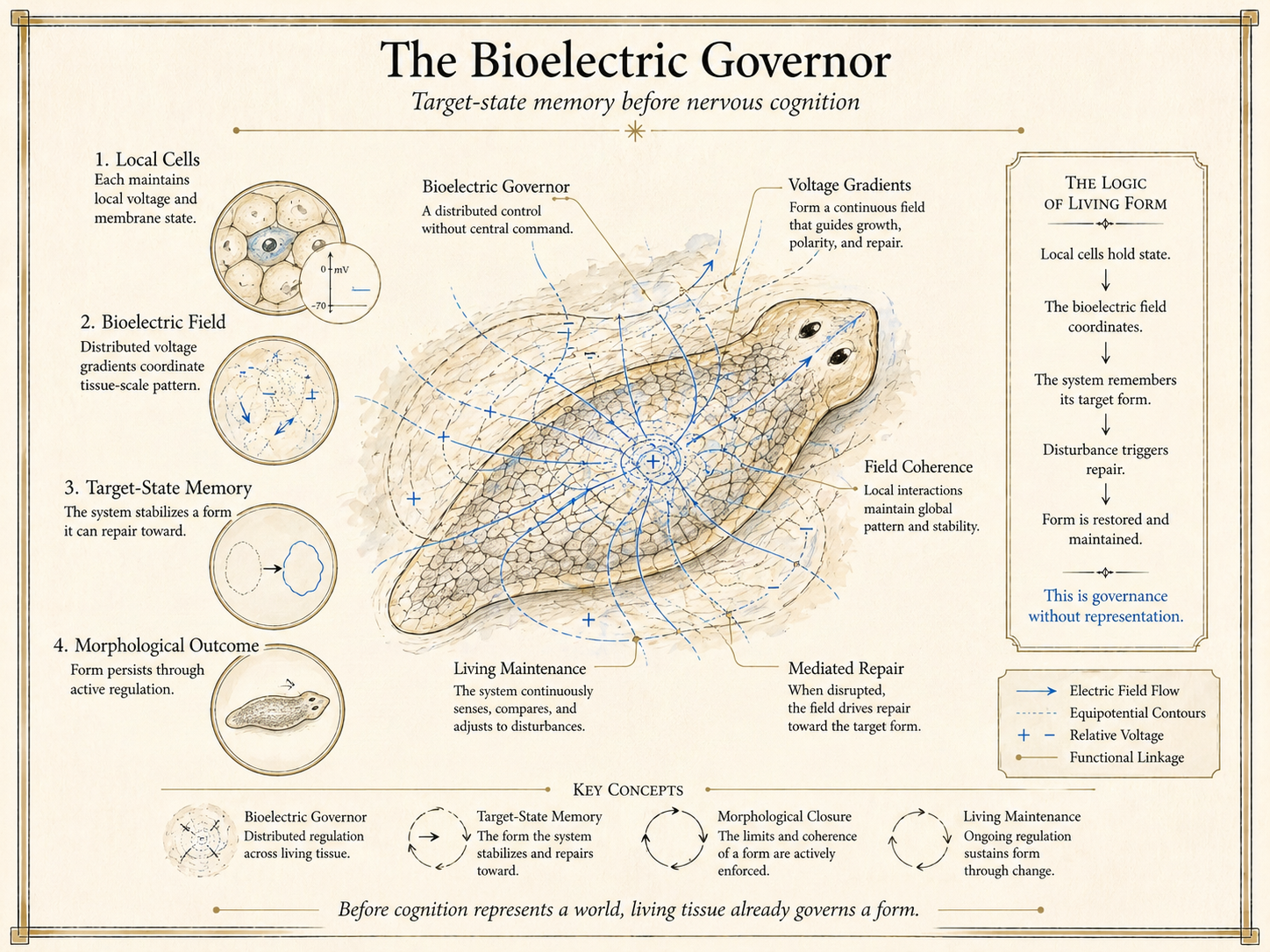
The bioelectric governor names a distributed regime of living form: local cells hold state, voltage gradients coordinate tissue-scale pattern, and the organism repairs toward a target morphology before nervous representation appears.
The one-sentence version
The genome is the parts list; the bioelectric governor is the construction plan. The Bioelectric Governor names the tissue-scale dynamical control variable — implemented through gap-junction-coupled transmembrane voltage patterns — that governs which morphogenetic target the tissue will converge on, independently of which proteins are present.
Where the word comes from
The evidence is brute experimental fact. Block gap junctions in a bisected planarian with octanol and the posterior fragment regenerates a second head where a tail should be — a two-headed organism, anatomically coherent at both poles, viable, mobile, and feeding, produced under a fixed genome by altering nothing but the bioelectric coupling topology that interprets that genome as a construction plan. Impose a specific transmembrane-voltage signature on the flank tissue of a Xenopus laevis embryo and a functional ectopic eye forms where no eye belongs: lens, retina, axonal wiring. Same genome. Same developmental stage. Different bioelectric governance. Different target.
Chapter 8 introduces the Bioelectric Governor as the first cognitive closure — not the only one, and not the highest one, but the stratum at which a genuine second-order constraint architecture is installed over the molecular inventory. The burn rate currency is ATP consumed by ion pumps maintaining transmembrane voltage gradients across the gap-junction network. The Witness function is the voltage pattern itself — a distributed, environment-redundant inscription of the tissue’s current constraint state. The Canon function is the attractor dynamics that select which of the available bioelectric states is load-bearing for downstream developmental governance.
Within the book’s Chapter 8-11 cognitive sequence, this names a local position in that sequence; the page’s R-register marks its broader placement in the site’s macro-map.
Why it matters
The Bioelectric Governor demonstrates, at its cleanest and most pharmacologically dissectible, the structure that every subsequent cognitive stratum will elaborate. Three features make it the framework’s paradigm case.
A control variable operates above the parts list. The molecular inventory specifies what is achievable; the bioelectric governor specifies what is selected from among the achievable. The two are distinct levels of description, and the bioelectric level is not reducible to the molecular one: the same molecular inventory generates radically different morphological outcomes under different bioelectric regimes.
The governance is operational without being mentalist. The tissue exhibits normativity — convergence on a target, correction of deviations from the target, characteristic failure modes when the governance architecture is perturbed — without any requirement for subjective experience, intention, or representation in any philosophically loaded sense. This is Chapter 8’s critical negative claim: operational normativity without interiority. The claim matters because it pre-empts both the temptation to project consciousness downward into tissue-level coordination and the temptation to dismiss tissue-level coordination as “merely mechanical.”
The failure modes are diagnostically specific. Attractor hijacking (the bioelectric landscape is rewritten toward a target that is coherent but wrong), gap-junction decoupling (the tissue loses the integration required for global target stabilisation), metabolic cessation (the ion pumps stop paying and the voltage gradients collapse), and plateau (the tissue stabilises without installing the governance capacity that would make the next stratum possible) — each failure mode is empirically distinguishable and mechanistically specific.
What it is not
The Bioelectric Governor is the first cognitive closure, not the cognitive stratum as a whole. Chapter 8’s central warning is that the cognitive stratum is a building with six floors, and the Bioelectric Governor is the ground floor. Most lineages stabilise here and go no further — the jellyfish is not a failed vertebrate, but the neural analogue of the prokaryotic plateau. What the Bioelectric Governor’s success generates is the reafference problem: a tissue-scale morphogenetic controller cannot distinguish self-generated perturbation from external perturbation at the speed and scale that locomotion demands.